Overview¶
The IV-HSL Emitter module produces and releases N-isovaleryl-L-homoserine lactone (IV-HSL), a branched acyl-homoserine lactone, enabling enzymatic small-molecule production, molecule release as a reporter output, inter-cell communication, and co-culture of synthetic cells with living bacteria. The module is based on work by Smith, Hartmann, and Booth, 2023.
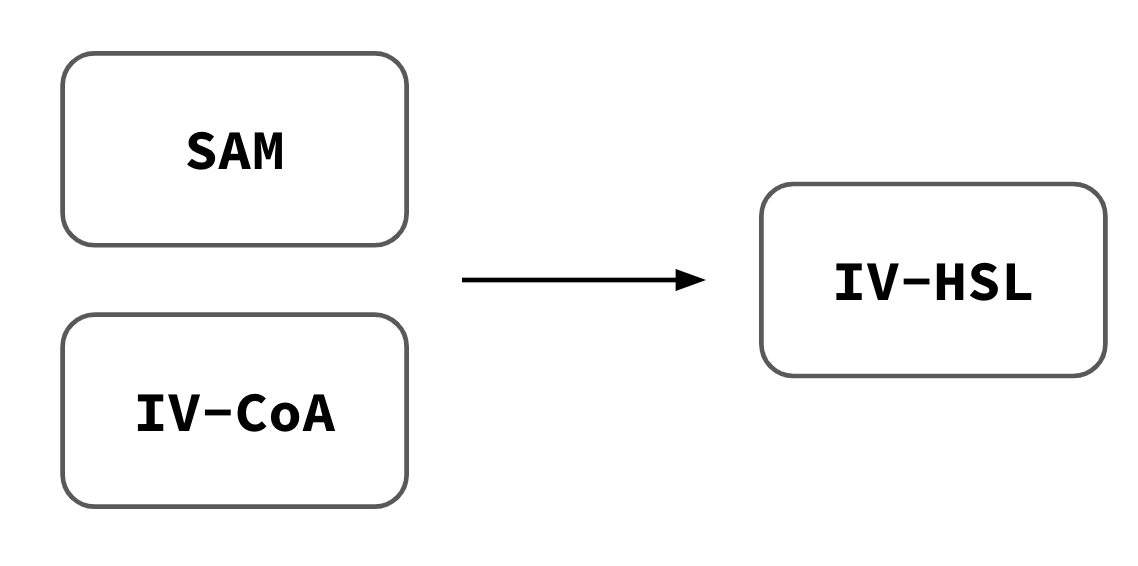
IV-HSL offers several advantages: it crosses synthetic cell membranes; its uncommon branched-chain structure makes it orthogonal from many other HSLs (Lindemann et al., 2011); and it activates expression in receiver cells at picomolar concentrations. The module encodes the BjaI enzyme under a constitutive T7 promoter. BjaI converts S-adenosylmethionine (SAM) and isovaleryl coenzyme A (IV-CoA) into IV-HSL, which diffuses out of the cell through the lipid bilayer. A companion E. coli receiver construct (bjaR-GFP-native) detects IV-HSL and produces a fluorescent output.
Design schematic of the IV-HSL Emitter module. pT7-bjaI expresses the BjaI enzyme, which converts SAM and IV-CoA substrates into IV-HSL. IV-HSL diffuses across the lipid bilayer and activates GFP expression in E. coli receiver cells via the BjaR transcription factor.
| Name | Length (bp) | File |
|---|---|---|
pOpen-pT7-bjaI | 2752 | pOpen-pT7-bjaI.gb |
pOpen-bjaR-GFP-native | 3877 | pOpen |
Cytosols¶
Usage¶
Add equimolar amounts of SAM and IV-CoA at 0.3 µM and 0.08 µM final concentration, respectively, to a standard PURE reaction containing pT7-bjaI. Volumes in µL.
| Component | Sample | Negative Control | Positive Control | Notes |
|---|---|---|---|---|
| PURE Solution A | 12 | 12 | 0 | Energy solution: small molecules |
| PURE Solution B | 9 | 9 | 0 | Proteins and ribosomes |
| RNase Inhibitor | 1.5 | 1.5 | 0 | Prevents RNase activity |
pOpen-pT7-bjaI (~200 ng/µL) | 1.5 | 0 | 0 | DNA encoding BjaI |
| SAM (5 mM) | 1.8 | 1.8 | 0 | Substrate for IV-HSL production |
| IV-CoA (5 mM) | 0.48 | 0.48 | 0 | Substrate for IV-HSL production |
| OptiPrep | 1.5 | 1.5 | 1.5 | Adds density for phase-transfer |
| IV-HSL (10 µM) | 0 | 0 | 0.3 | Commercial IV-HSL for positive control |
| 3M Glucose | 0 | 0 | 8.46 | |
| ddH₂O | 2.22 | 3.72 | 19.74 | |
| Total | 30 | 30 | 30 |
Expected Performance¶
PURE reactions were incubated for 4 hours, then added to log-phase E. coli receiver cell cultures containing bjaR-GFP-native. GFP fluorescence was measured over 6 hours at 5-minute timepoints using a BioTek Cytation 5 plate reader. Expression in XL-10 Gold cells containing bjaR-GFP-native is equivalent for bulk PURE reactions with pT7-bjaI and substrates and for emitter cells containing IV-HSL without plasmid DNA — both show a significant response over the negative control.
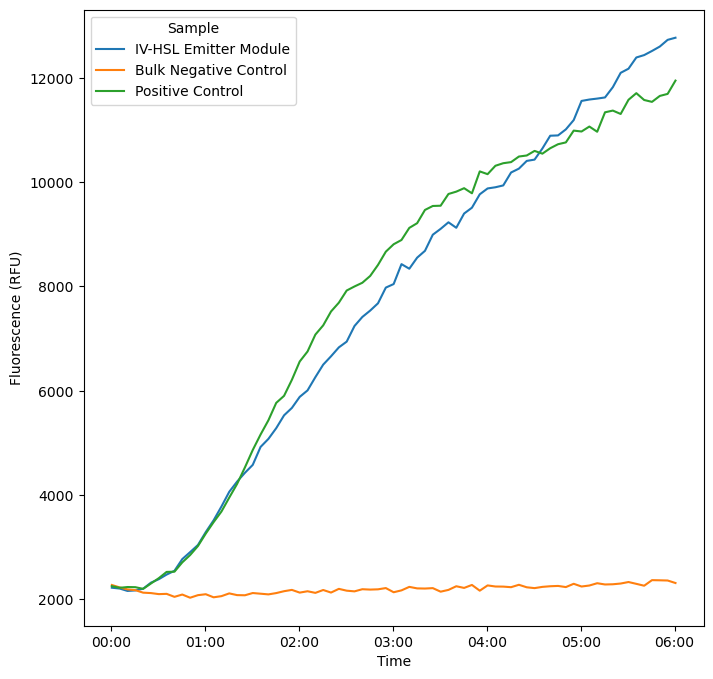
Expression kinetics of E. coli receiver cells modified with bjaR-GFP-native in response to IV-HSL produced by PURE reactions.
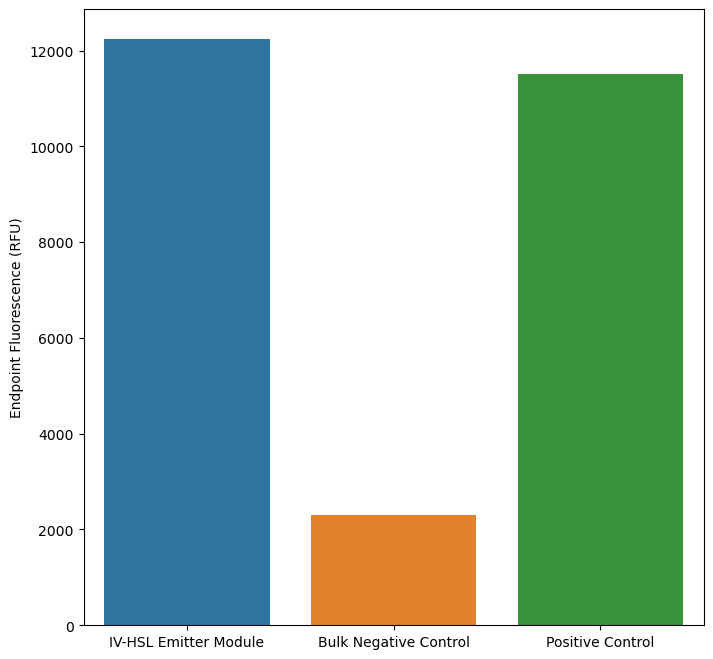
Expression of E. coli receiver cells modified with bjaR-GFP-native at steady state.
Cells¶
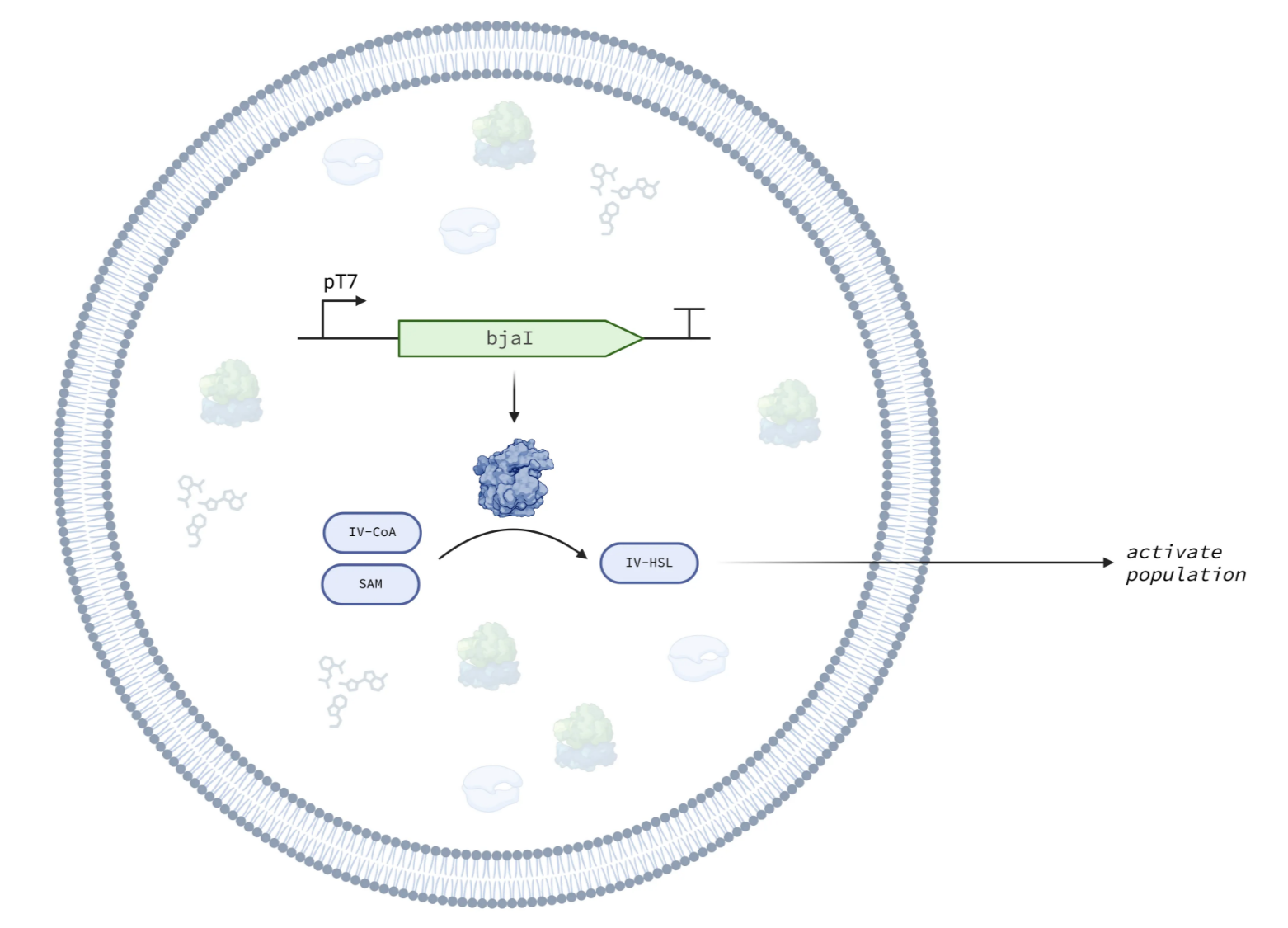
The IV-HSL Emitter module in the Base Cell.
Expected Performance¶
Emitter Cells were constructed and co-cultured with E. coli containing bjaR-GFP-native. Time-series confocal microscopy (Revvity Operetta CLS) was performed over 8 hours collecting red (Rhodamine-B), green (GFP), and brightfield images at 40× magnification across multiple fields per well at approximately 15-minute intervals.
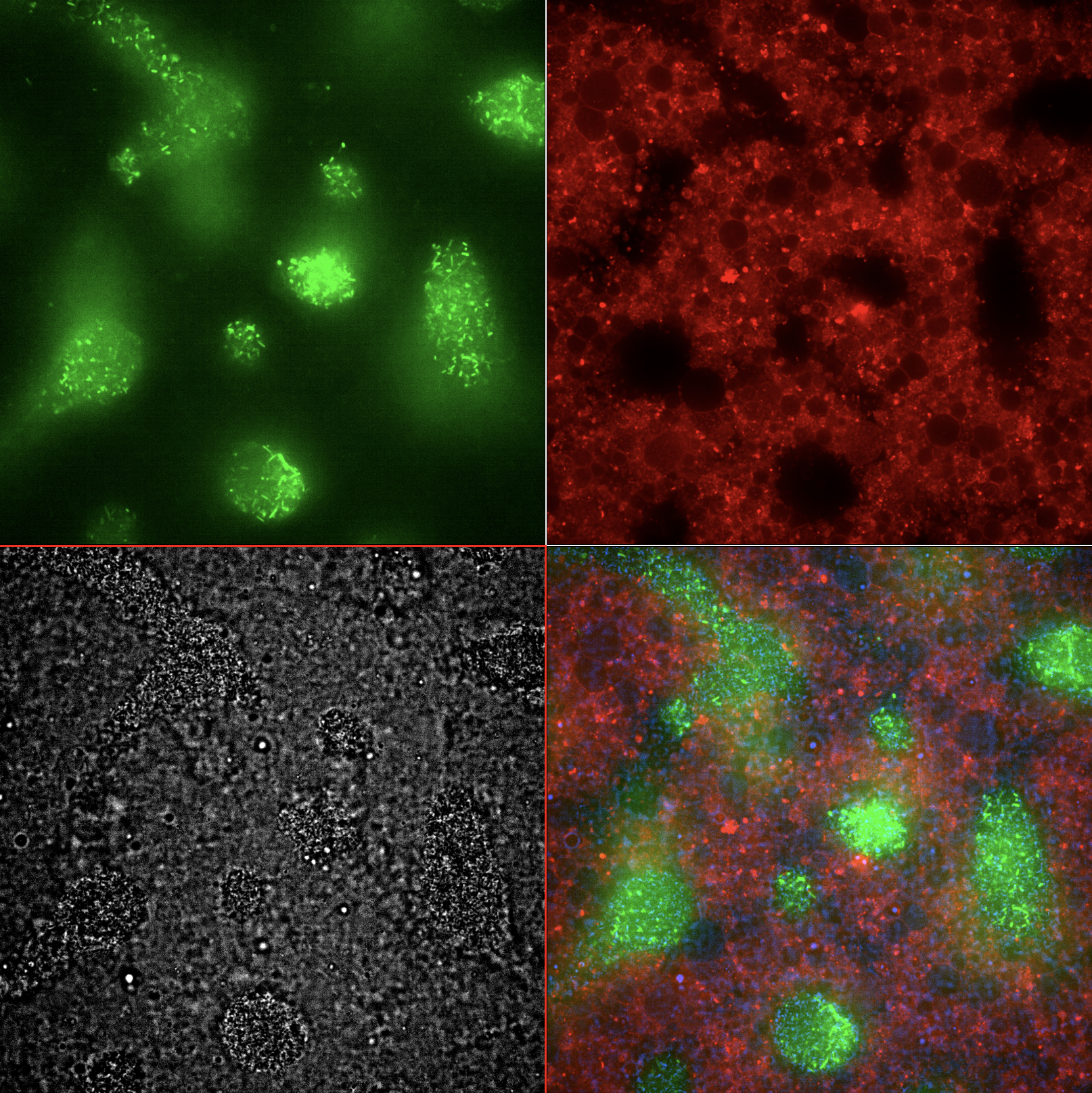
Emitter Cell Endpoint Montage. Single field of view at t = 8 hours. (green) E. coli producing GFP in response to IV-HSL emitted by the Emitter Cells. (red) Emitter cells with rhodamine-labeled membrane. (gray) Brightfield. (rgb) Merged image.
The Emitter Cell causes E. coli to express GFP in response to IV-HSL.
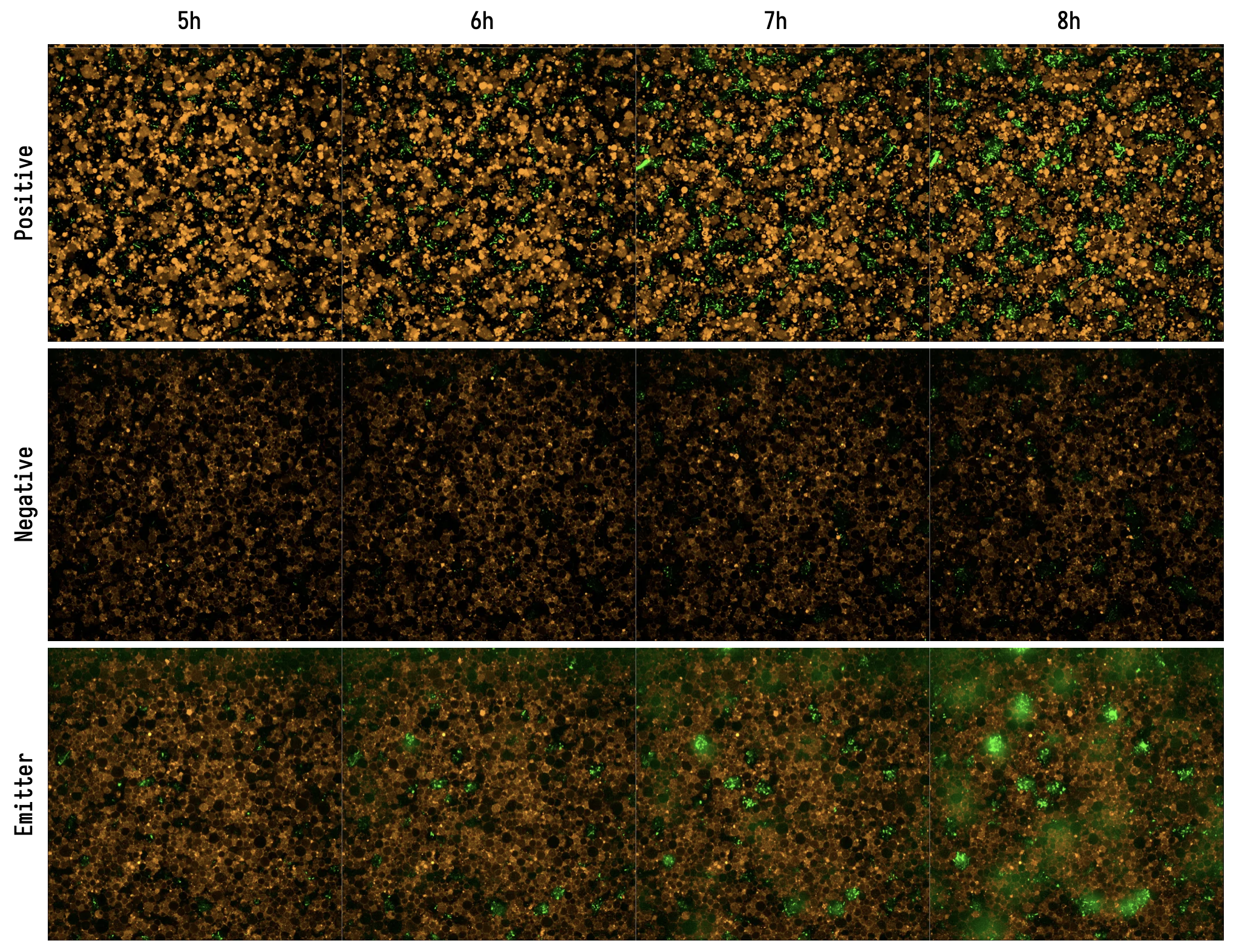
Emitter Cell Timeseries. (Positive) Liposomes contain PURE and 100 nM IV-HSL. (Negative) Liposomes contain PURE supplemented with SAM and IV-HSL, but no DNA encoding BjaI. (Emitter) Liposomes contain PURE expressing BjaI from pT7-bjaI. Exposures are matched between wells. Each field of view is 167 µm wide.
- Smith, J. M., Hartmann, D., & Booth, M. J. (2023). Engineering cellular communication between light-activated synthetic cells and bacteria. Nature Chemical Biology, 19(9), 1138–1146. 10.1038/s41589-023-01374-7
- Lindemann, A., Pessi, G., Schaefer, A. L., Mattmann, M. E., Christensen, Q. H., Kessler, A., Hennecke, H., Blackwell, H. E., Greenberg, E. P., & Harwood, C. S. (2011). Isovaleryl-homoserine lactone, an unusual branched-chain quorum-sensing signal from the soybean symbiont Bradyrhizobium japonicum. Proceedings of the National Academy of Sciences, 108(40), 16765–16770. 10.1073/pnas.1114125108